精子・卵子を形成する上で鍵となる染色体構造を発見
染色体の担い手「コヒーシン蛋白質」が支える染色体のしなやかさが重要!
| 原著論文 | Nucleic Acids Res. gkac183 (2022) |
|---|---|
| 論文タイトル | Rec8 cohesin-mediated axis-loop chromatin architecture is required for meiotic recombination |
| 研究室サイト | 細胞核ダイナミクス研究グループ〈平岡 泰 招へい教授〉 |
概要
大阪大学大学院生命機能研究科の作野剛士特任准教授(常勤)と平岡泰教授らの研究グループは、減数分裂期における組換えを開始するためには、コヒーシンによって作られる軸状染色体構造の形成が必要であることを世界で初めて明らかにしました。
遺伝情報を次世代に伝えるためには、減数分裂により精子や卵子が作られる必要があります。減数分裂ではまず染色体DNAが複製された後に、両親由来の染色体の間で組換え反応が起こります。複製の際、元のDNAと新たなDNAを繋ぎ留めておく「接着因子」がコヒーシンです。コヒーシンは接着に加えて、軸状染色体構造の形成や組換え反応に必要な因子を染色体へと呼び込む上でも機能していることが知られていました(図1)。しかし軸状染色体構造の意義や、その構造と組換え反応との関連性についてはこれまで明らかになっていませんでした。
今回、分裂酵母をモデルとして解析を行った結果、減数分裂期組換えの前におこる染色体のペアリングに、軸状染色体構造が必要であることがわかりました。さらに、軸状染色体構造を形成できなくなるコヒーシン変異体では、減数分裂期組換えの開始に必要な因子群が染色体に集まれなくなり、組換え反応に著しい欠損を示すことも明らかになりました(図2)。ヒトでは、減数分裂期の組換えにエラーが生じると、ダウン症や不妊につながると考えられています。コヒーシンや軸状染色体構造はヒトにも存在することから、今回明らかになった知見は将来的に不妊治療の進展に寄与することが期待されます。
研究の背景
我々ヒトをはじめ多くの生物は、精子や卵子などの配偶子に両親の染色体を半分ずつ分配し、受精により組み合わせることで遺伝情報を次の世代に伝えます。配偶子は、減数分裂という特殊な染色体を分配する仕組みによって生殖細胞から形成されます。減数分裂過程では、両親由来の染色体がペアリングし、その間で組換えと呼ばれる反応が起こります。その結果、互いのDNA情報を混ぜ合わせると同時に互いの染色体を結びつけることになり、たくさんある染色体の中で分配する組み合わせを間違えないようにする仕組みが用いられています。しかしこの組換え反応がどうやって開始されるのか、そのメカニズムはよく分かっていませんでした。ヒトでは組換えの過程にエラーが生じると、ダウン症や不妊につながると考えられています。よって、組換えを解析することで減数分裂のプロセスに関して正しい理解が進み、将来的には不妊治療や生殖医療の進展に寄与することが期待されます。
本研究の成果
今回、研究グループでは分裂酵母を用いて、減数分裂の際にコヒーシンによって形成される軸状染色体構造と組換え反応との関係について解析を行いました。その結果、染色体にコヒーシンが無い場合だけでなく、過剰にある場合についても染色体のペアリングがうまくいかなくなることが分かりました。ペアリングは、染色体の両端を核膜にくっつけて染色体全体を湾曲させながら揺り動かすことにより促進されますが、コヒーシンが無い場合は染色体がほとんど動かず、過剰にある場合は染色体の湾曲が観察されませんでした。染色体の構造を蛍光顕微鏡やHi-C法を用いて解析すると、コヒーシンが無い場合は軸状染色体構造が形成されず、コヒーシンが過剰にある場合は軸状染色体構造の軸部分が太くなる一方でDNAのループが長くなることが明らかになりました。よって、染色体全体をうまく動かしペアリングを促進するためには、コヒーシンが軸状染色体構造を適切に形成し、染色体の最適な「硬さ」を生み出すことによって実現されている可能性が示されました(図3)。
また、遺伝学的解析が駆使できる分裂酵母の特性を活かして、接着の機能は維持したまま軸状染色体構造が形成できなくなるコヒーシン変異体を見つけることに成功しました。この変異体では軸状染色体構造が無くなるので、予想通りペアリングが出来ませんでした。またこのコヒーシン変異体は染色体に存在するにもかかわらず、コヒーシンを無くした場合と同様に、組換えに必要な因子群が染色体上へ集積できないことが明らかになりました。それに伴って、組換え反応もほぼ起こらなくなることも判明しました。これらの結果から、減数分裂の際コヒーシンによって形成される軸状染色体構造の生物学的な意義がはじめて明らかになりました。すなわち、これらの特殊な染色体構造は、組換え因子を安定的に染色体へと呼び込む上での足場となり、組換え反応の実行を下支えしていたのです。
研究成果のポイント
- 精子や卵子の形成に必須な減数分裂における染色体のペアリングには染色体の適度な「しなやかさ」が必要
- 染色体の屋台骨、コヒーシンは「接着」と「染色体の形作り」という別々の機能を持つことを解明
- コヒーシンが支える染色体の「形」が減数分裂期の組換えを開始するために必要
- 減数分裂に対する理解が深まり、将来的に生殖医療の進展に寄与することが期待される
本研究成果が社会に与える影響(本研究成果の意義)
分裂酵母を用いた今回の解析により、組換え制御における新たな機能が明らかになったコヒーシンは、進化的に保存されていてヒトにも同様な因子が存在します。また、接着の機能は維持したまま、軸状染色体構造が形成できなくなるコヒーシン変異体に関して、その変異箇所周辺のアミノ酸配列がヒトにおいても保存されていることから、今回明らかになった「コヒーシンによって作られる軸状染色体構造が、組換え因子を安定的に染色体へと呼び込む上での足場になる」という機構も進化的に保存されている可能性が十分に考えられます。減数分裂期組換え反応に異常が生じると、染色体分配のエラーが誘発される結果、不妊やダウン症の要因となることが知られています。よって、今回の発見は、組換え反応を制御する根源的なメカニズムの解明につながる成果であり、ひいては組換えに依存した減数分裂期の染色体分配を制御する仕組みの理解につながり、将来的には不妊治療などへの応用が期待されます。
研究者のコメント
「軸構造とDNAループからなる特殊な染色体構造」が減数分裂期に形成されるという事実は、約100年も前から顕微鏡観察によって明らかになっていました。しかし、これまでその構造の生物学的な意義に関しては謎のままだったのですが、今回の一連の解析からその謎の一端を明らかにすることが出来ました。今後は、ではどうやってコヒーシンがそのような染色体構造を作り出すのか、そのメカニズムを明らかにするべく研究を行っていきます。(作野剛士)
特記事項
本研究結果は、イギリスの科学雑誌「Nucleic Acids Research」において、3月25日(金)に公開されました。
なお本研究は、日本学術振興会科学研究費助成事業若手研究A「減数第一分裂期における染色体分配制御機構の解析(研究代表者:作野剛士)」、新学術領域研究「減数分裂における細胞核・クロマチン構造の変換メカニズム(研究代表者:平岡泰)」、アメリカ国立衛生研究所研究費R01(研究代表者:野間健一)の一環として行われました。
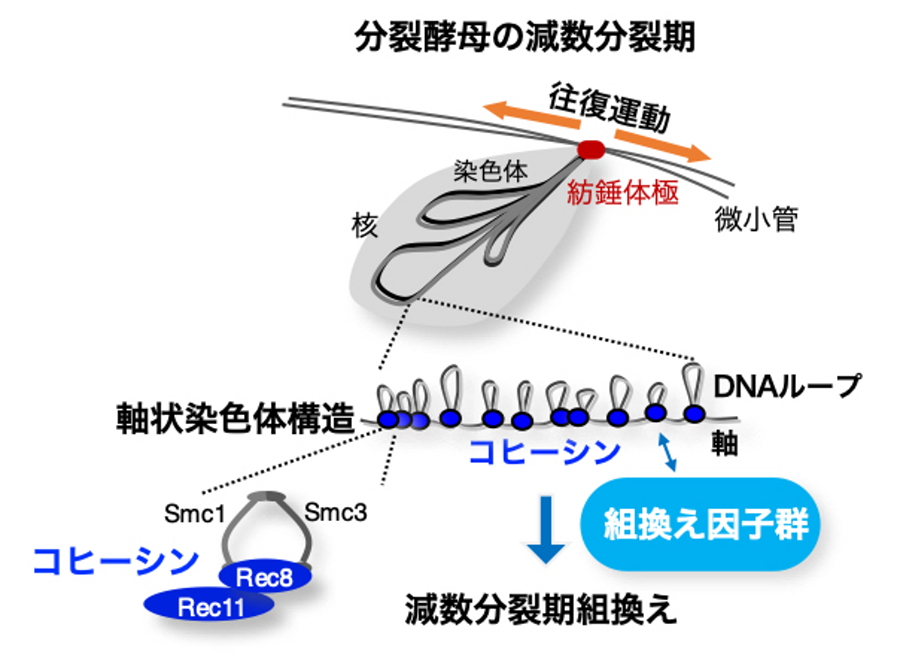
図1.分裂酵母の減数分裂期における染色体の模式図
染色体の両末端が紡錘体極で束ねられた状態で、紡錘体極を起点に核が細胞内を往復運動することで染色体のペアリングが促進される。左下に示すコヒーシンは複合体で、軸上染色体構造形成の寄与に加え、染色体相同組み換えの因子を呼び込む機能がある。
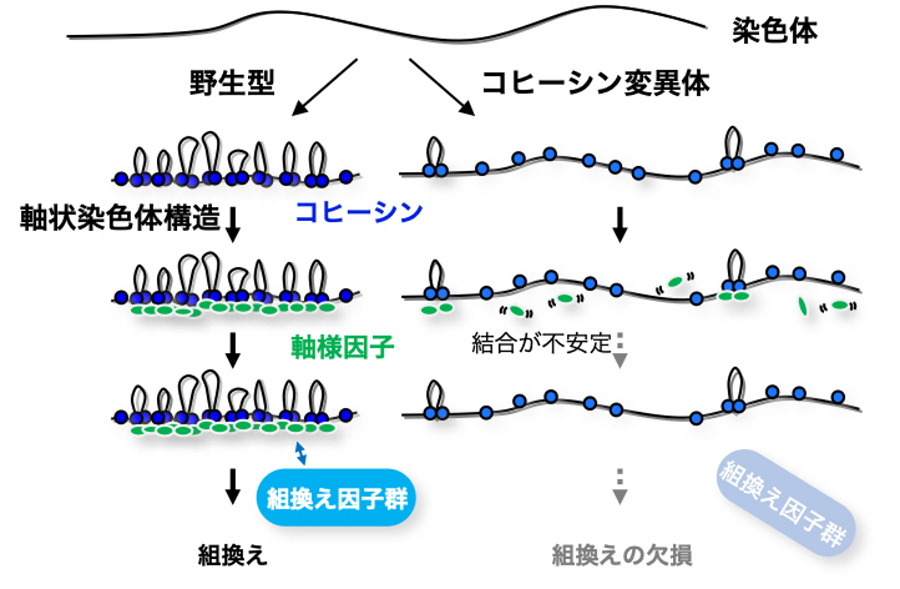
図2.減数分裂期の組換えに必要な染色体における染色体の模式図
軸様因子はコヒーシンが作る軸状染色体構造に依存して染色体上に安定的に形成される。軸様因子は組換え因子群を染色体へ呼び込むことで、組換えが実行される。右側の軸状染色体構造が形成できないコヒーシン変異体では、軸様因子と染色体の結合が不安定になる結果、組換え因子が染色体に呼び込まれないので組換えがおこらない。
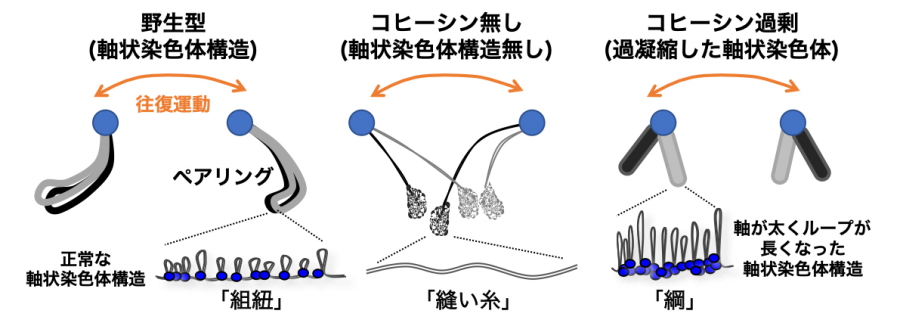
図3.減数分裂期における染色体の動きを模式的に示した図
減数分裂期では、末端のテロメア(青丸)を核膜に結合した状態で往復運動する。その間に染色体は「組紐」のようにしなやかに動いて湾曲することでペアリングする(野生型)。コヒーシンが無い、または軸状染色体が形成されないと、染色体は細い「縫い糸」のようになってしまい、テロメアの動きが染色体全体に伝わらずにペアリングしない。一方で、コヒーシンが過剰になると染色体は「綱」のように硬い構造体へと変化し、往復運動の動きに合わせた染色体の湾曲が起こらなくなりペアリングしない。
用語解説
- 減数分裂
生殖に必要な配偶子(精子や卵子など)を形成する際に行われ、生じた配偶子では染色体数が分裂前の細胞の半分になる。 - 染色体
遺伝情報を担うDNAとタンパク質の構造体。ヒトの細胞では、父親と母親に由来する23組46本の染色体をもつことが知られている。 - ペアリング
両親由来の染色体ペアの間で組換えを行うために、組換えに先立って正しい相手を認識して並び合う現象で、減数分裂期にのみ観察される。 - コヒーシン
リング状の形状をした4つの蛋白質からなる複合体で、リングの穴に2本の染色体DNAを通すことで、それらの接着を担うと考えられている(図1参照)。 - 減数分裂期の組換え
両親由来の染色体ペアの間でDNA鎖を繋ぎ変えて交換する反応で、結果的に染色体ペアが互いにつながることになる。 - 軸状染色体構造
染色体上のコヒーシンが軸状に集まると同時に、一定距離間隔でのDNAループ形成を伴った構造体で、結果として染色体全体がコンパクトになる(図1参照)。また、この構造の形成にはコヒーシンが必要である。 - 分裂酵母
ヒトと同じ真核生物に属する単細胞生物で、自己の増殖や減数分裂という生き物としての根幹に関わるメカニズムの多くはヒトと分裂酵母の間で保存されている。 - ダウン症
減数分裂の異変に起因して、21番染色体を一本余分に受け継ぐことにより生じる病気。 - 生殖細胞
減数分裂を行い、精子や卵子を作るために特化した細胞の集まり。 - Hi-C法
high–throughput chromosome conformation captureの略語で、細胞を観察したい条件で生きたまま固定して、DNAの立体構造を保ち、DNAを断片化した後にDNA断片同士をつなぐことで、立体構造の内部で近接していたDNA領域を網羅的に捉え、染色体DNAの三次元的な空間配置を知るための実験手法。
| 原著論文 | Nucleic Acids Res. gkac183 (2022) |
|---|---|
| 論文タイトル | Rec8 cohesin-mediated axis-loop chromatin architecture is required for meiotic recombination |
| 著者 | Takeshi Sakuno (1), Sanki Tashiro (2), Hideki Tanizawa (2), Osamu Iwasaki (2), Da-Qiao Ding (3), Tokuko Haraguchi (1), Ken-Ichi Noma (2, 4), Yasushi Hiraoka (1)
|
| PubMed | 35333350 |