いのちの始まりを導くエピジェネティックスイッチ
ヒストン修飾H3K9me2の動的制御が遺伝子発現の目覚めを促す
| 原著論文 | Nat. Struct. Mol. Biol. 33(6): 928-938 (2026) |
|---|---|
| 論文タイトル | Dynamic regulation of H3K9 di-methylation drives mouse minor zygotic genome activation |
| 研究室サイト | エピゲノムダイナミクス研究室〈立花 誠 教授〉 |
概要
大阪大学大学院生命機能研究科の前田亮助教、黒木俊介准教授、立花誠教授、理化学研究所生命医科学研究センター疾患エピゲノム遺伝研究チームの井上梓チームディレクターらの研究グループは、受精後の初期胚が自身のゲノムを使い始める「胚性ゲノム活性化(ZGA)」の精密な制御機構を解明しました。特にその第一波である「マイナーZGA」において、ヒストン修飾H3K9me2が受精卵(2細胞期初期)のマイナーZGA遺伝子領域から一時的に消失し、その後回復するという動的な変化を発見しました。
この消失には卵子に蓄えられたH3K9脱メチル化酵素KDM3A/Bが必須であり、欠損するとマイナーZGA遺伝子の発現低下と発生停止が起きることを示しました。さらに、マイナーZGA遺伝子の抑制にはH3K9メチル化酵素SETDB1とG9aが協調的に機能することを明らかにしました(図1)。
本研究成果は、不妊治療やリプログラミング技術の発展への貢献が期待されます。
研究の背景
受精直後の胚は自身のゲノムからの転写を停止しており、卵子が蓄えた母性因子(mRNAやタンパク質)によって発生が制御されています。しかし、母性因子は受精後次第に分解されるため、胚は自身のゲノムから転写を開始しなければなりません。この胚由来の遺伝子発現の開始を「胚性ゲノム活性化(ZGA)」と呼びます。
ZGAは段階的に進み、マウスでは1細胞期後半から2細胞期にかけて起きる第一波を「マイナーZGA」と呼びます。この時期には、他の細胞種では発現が見られない特異的な遺伝子やレトロトランスポゾンが一過的に活性化され、全能性の獲得に重要な役割を果たすと考えられています。しかし、これらのマイナーZGA遺伝子の発現がこの時期に精密に活性化・抑制される分子機構は未解明でした。
また、ゲノムDNAは「ヒストン」とよばれるタンパク質に巻き付いており、アセチル化やメチル化などヒストンへの化学修飾が転写活性を調節することが知られています。H3K9me2(ヒストンH3の9番目のリジンのジメチル化)は遺伝子発現が抑制されたゲノム領域にみられる代表的なエピジェネティックマークです。この修飾は、H3K9メチル化酵素によって付加され、脱メチル化酵素によって除去されることで、細胞内では動的な平衡状態を保っています。これまでの研究から、H3K9me2が2細胞期前後に全体的に低下することは報告されていましたが、この変動がZGAの制御とどのように関係するかは不明でした。
本研究の成果
1. マイナーZGA遺伝子領域のH3K9me2は2細胞期あたりにおいて一過的に消失する
研究グループはまず、卵母細胞・2細胞期初期(E2C期)・桑実胚期のH3K9me2のゲノムワイドな分布を解析しました。その結果、マイナーZGA遺伝子領域周辺のH3K9me2がE2C期に一過的に消失することが明らかになりました(図2)。この低下は、RNAポリメラーゼIIの結合増加と逆相関しており、H3K9me2の消失が転写活性化と対応していることが示されました。H3K9me3やH3K27me3といった他の抑制性ヒストン修飾にはこのような変化は見られず、H3K9me2に特異的な現象でした。これらのことから、「H3K9me2の減少がマイナーZGAの発現を引き起こす」のではないかと考えました。
2. 母性因子KDM3A/BがマイナーZGAを駆動する
次に研究グループは、「H3K9me2メチル化・脱メチル化のバランスの変化」がE2C期にH3K9me2の消失をもたらす原因ではないかと考えました。この仮説を検証するため、Kdm3a/bを卵子で欠損させた受精卵を作製したところ、大多数が2〜4細胞期で発生停止することが分かりました。この欠損胚では、H3K9me2が増加し、マイナーZGA遺伝子の約6割が有意に発現低下していました(図3)。このマイナーZGA遺伝子の発現低下は、Kdm3aのmRNAを受精卵に戻すことにより部分的に回復しましたが、酵素活性のない変異体では回復しませんでした。これらの結果から、母性KDM3A/BによるH3K9me2の脱メチル化が、マイナーZGAに必要であることが証明されました。
3. SETDB1とG9aは協調的にマイナーZGA遺伝子の発現を抑制する
マイナーZGA遺伝子は2細胞期を過ぎると急速に抑制されます。mESCを用いた解析から、H3K9メチル化酵素SETDB1とG9aを同時に欠損させると発現が相乗的に上昇することを見出しました。マイナーZGA遺伝子は、DUXという転写因子によって活性化されることが知られています。SETDB1はDuxの発現を特異的に抑制し、G9aはマイナーZGA遺伝子領域の広範囲にわたってH3K9me2を付加することで、これらの遺伝子を核膜付近に局在させ遺伝子発現を抑制することが明らかになりました。すなわち、両酵素は異なる機構を通じて協調的にマイナーZGA遺伝子の再活性化を防いでいることが示されました。
研究成果のポイント
- 2細胞期初期の受精卵において、遺伝子の発現を抑制する「スイッチ」となるヒストン修飾H3K9me2が、受精直後に発現する遺伝子(マイナーZGA遺伝子)領域から一時的に消失することを発見。
- H3K9me2を除去する酵素KDM3A/Bが失われるとマイナーZGA遺伝子の発現が大きく低下し、受精卵の発生が停止することが分かった。また、マイナーZGA遺伝子の抑制には別の酵素SETDB1とG9aが協調して働いていることが明らかに。
- 受精という「いのちの始まり」の瞬間における、ゲノムを眠りから覚ます「エピジェネティックスイッチ」の精密な制御の様子が明らかになったことから、クローン作製(体細胞核移植)やiPS細胞作製における「リプログラミング」技術の効率改善、不妊治療の新たなアプローチへつながることに期待。
本研究成果が社会に与える影響(本研究成果の意義)
受精という「いのちの始まり」の瞬間、ゲノムを眠りから覚ます「エピジェネティックスイッチ」が精密に制御されていることを、今回の研究が明らかにしました。この知見は、発生生物学の基礎的な問いに答えるだけでなく、応用面でも重要な意味を持ちます。
H3K9me2はクローン作製(体細胞核移植)やiPS細胞作製におけるリプログラミングの障壁としても知られており、その制御機構の解明は、これらの技術の効率改善につながることが期待されます。また、初期胚の発生停止は不妊の一因でもあり、本研究で示された分子機構は不妊治療の新たなアプローチを探る上でも基盤となる知見です。
研究者のコメント
受精卵がどのようにして自分の遺伝子を使い始めるのか、その仕組みは発生生物学における根本的な問いの一つです。今回、H3K9me2という一つのヒストン修飾の動的な変化が、マイナーZGA遺伝子の活性化と抑制に重要な役割を果たしていることを明らかにできました。この発見が、不妊治療やリプログラミング技術の発展につながることを期待しています。(前田 亮)
特記事項
本研究成果は、2026年5月26日(火)18時(日本時間)に英国科学誌「Nature Structural and Molecular Biology」(オンライン)に公開されました。
本研究は、JSPS科研費(26H01528、26H01056、24H02320、24H00556、24K09326、24H01368、23K18095、22H04671)の支援を受けて実施されました。
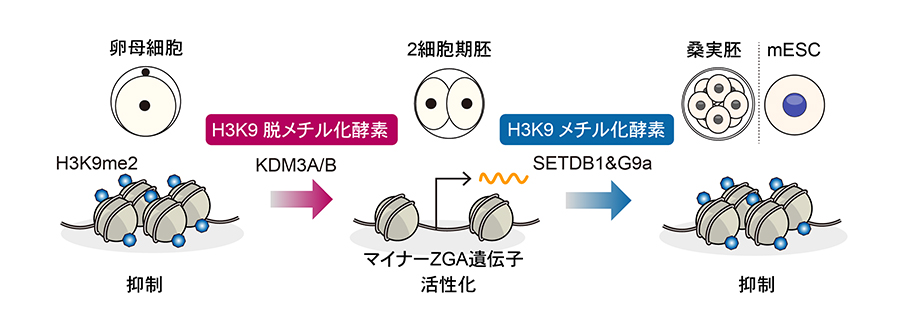
図1. H3K9me2の動的変化によるマイナーZGA遺伝子の制御
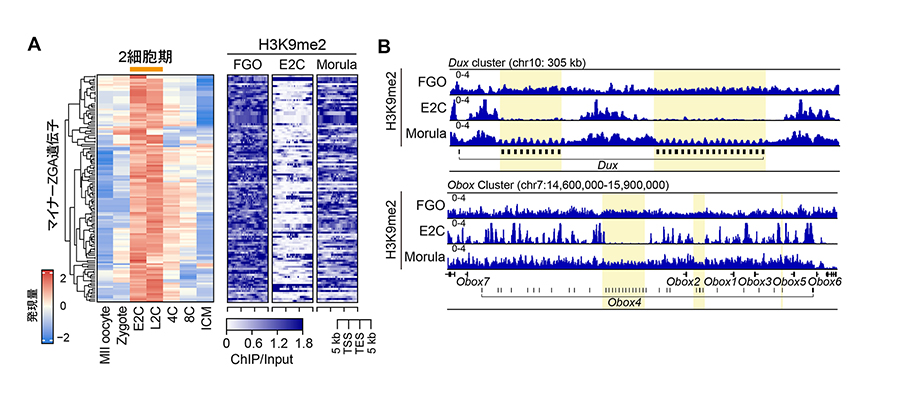
図2. 初期胚におけるマイナーZGA遺伝子の発現とH3K9me2プロファイル
A. 左) マイナーZGA遺伝子は2細胞期をピークに発現する。右) マイナーZGA遺伝子上のH3K9me2は2細胞期初期(E2C)で消失する。FGO、卵母細胞; Morula、桑実胚。
遺伝子発現データはWu et al (2016), FGOのH3K9me2データはAu Yeung et al (2019)より取得。
B. 代表的なマイナーZGA遺伝子上のH3K9me2の変化。DUXやOBOX4(黄色部分にコード)はZGAを駆動する転写因子である。
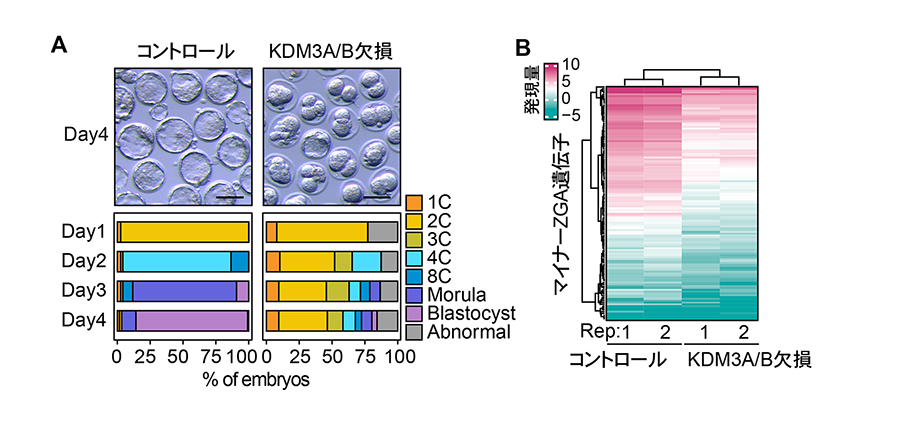
図3. KDM3A/B欠損が初期発生とマイナーZGA遺伝子発現に与える影響
A. KDM3A/B欠損胚の多くは2細胞期(2C)から4細胞期(4C)の間に発生が停止する。
B. 2細胞期初期胚において、KDM3A/B欠損によりマイナーZGA遺伝子の発現が低下する。
用語解説
- 転写
DNAの遺伝情報をもとにRNAを合成する過程。遺伝子が「使われている」状態を指す。 - レトロトランスポゾン
ゲノム内に多数コピーが存在する反復配列の一種。通常は抑制されているが、初期発生など特定の時期に活性化し、遺伝子発現の調節に関与する。 - 全能性
一つの細胞がからだのすべての細胞種(胎児本体および胎盤など胚外組織を含む)に分化できる能力。受精卵や初期胚の細胞に備わる。 - エピジェネティックマーク
DNAの塩基配列を変えずに遺伝子の働きを調節する化学的な印。ヒストンへのメチル化・アセチル化などが代表例。 - RNAポリメラーゼII
遺伝子のDNAを読み取りmRNAを合成する酵素。この酵素が遺伝子領域に結合しているほど、その遺伝子が活発に転写されていることを示す。 - 卵子で欠損させた受精卵
受精直後の胚は卵子から持ち込まれた母性因子によって制御されている。母性因子としての機能を調べる方法の一つとして、卵子特異的に目的タンパク質を欠損させる手法がある。 - 核膜付近
核ラミナ。細胞核の内側を裏打ちする構造。この領域に局在する遺伝子は転写が抑制される傾向がある。
| 原著論文 | Nat. Struct. Mol. Biol. 33(6): 928-938 (2026) |
|---|---|
| 論文タイトル | Dynamic regulation of H3K9 di-methylation drives mouse minor zygotic genome activation |
| 著者 | Ryo Maeda (1, 6), Shunsuke Kuroki (1, 6), Hiromi Shimojo (2), Masahiro Nagano (3, 4), Masahiro Matsuwaka (5), Hiroshi Sasaki (2), Azusa Inoue (5), Makoto Tachibana (1)
|
| PubMed | 42191875 |