


4. 多繊毛同調運動におけるTJ-アピカル複合体の役割
気管多繊毛上皮細胞におけるTJ-アピカル複合体の役割
多繊毛上皮細胞の運動性繊毛は、細胞内・細胞間で協調しており、気管では肺側から口側への粘液流を作り感染源を排泄するために、卵管では卵子および受精卵を輸送するために、脳室上衣では脳脊髄液流を作り出すために必須です。発生期におけるノード領域の繊毛運動は身体の左右軸決定に深く関わっていることが知られています。このように運動性繊毛の発生及び生体恒常性維持への貢献は大きく、繊毛運動のメカニズムについての研究は盛んに行われています。しかしながら、多繊毛が細胞内・細胞間でいかに協調性を獲得するかは不明な点が多く、多繊毛協調運動の制御機構の解明は喫緊の課題と認識されています。本研究では、多繊毛協調運動と「TJ-アピカル複合体」を関連付けて解析を進めており、これにより上皮細胞機能研究における新たな視点が開けるものと考えられます。
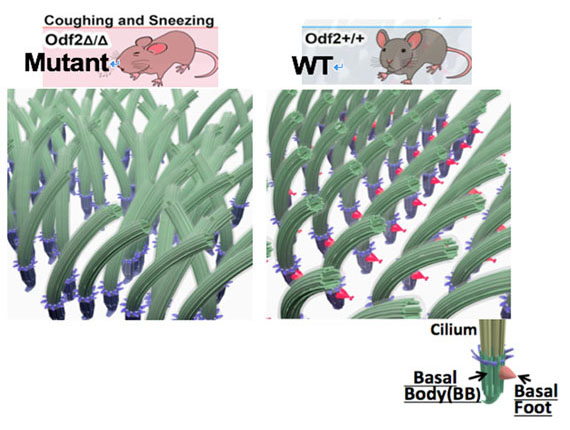
図1.Odf2KOマウスにおける多絨毛配列様式。
BFにより多絨毛の配列パターンが決まるため、Odf2KOによりBFが欠失すると、絨毛配列が大幅に乱れ、咳などの症状が現れます。
繊毛の根元には、中心体に由来する基底小体(BB: Basal Body) という骨格構造体があり、繊毛運動の方向性を決めています(図1)。BBにはAnchoring fiber(AF),Basal foot(BF)という2種類の突起構造があり、AFは基底小体と細胞膜との結合に、BFは基底小体の方向決定に関わることがわかっています。当研究室で同定したBB関連蛋白質であるOdf2は、AF,BFの構成タンパク質 として知られている数少ない蛋白のひとつです。このOdf2変異マウスにおいて、咳とくしゃみの表現型がみられ、気管上皮における多繊毛協調運動の欠失が確認されました(図1)。さらに超高圧電子顕微鏡トモグラフィー(UHVEMT)による検証から、BFが欠失し、野生型で均一なBBの空間配置が乱れていることが明らかになっています。また検証の過程で、野生型の気管多繊毛上皮表層には、格子状に微小管が分布し、各格子に1つずつのBBが存在するのに対し、Odf2変異マウスでは微小管格子が欠失することが明らかになりました。これにより、BF形成にはOdf2が必須であること、気管上皮表層の微小管格子とBFが密接に関係して多繊毛協調運動が創出されることが明らかになりました。しかしながら、そのメカニズムは未だ不明です。

図2.気管多絨毛上皮細胞における絨毛根元の基底小体(BB)配列の規則性の確立過程の長時間高解像ライブイメージング。
A.GFP-セントリンでラベルしたBBのライブイメージングと各ステージのBBの配向性をOdf2(緑)とcentriolin(赤)のラベルで示しています。
B.4つの配列様式においての解析結果が以下の方法で示されました。1.Odf2の蛍光色、2.超高圧電子顕微鏡トモグラフィー(UHVEMT)での微小管、3.数理生物学的シミュレーション。
現在、「TJ-アピカル複合体」の視点から、多繊毛運動の協調性獲得メカニズムについて検討を進めています。
長時間高解像ライブイメージングによる多繊毛上皮細胞の繊毛の規則的配列構築メカニズムの解明
私共では、繊毛根元のBBの規則的配列過程について、世界で初めての長時間高解像ライブイメージングを開発して、その過程をリアルタイムで撮影することにより、その原理を明らかにしました。BBをGFP-セントリンでラベルしたマウスから調製した気管上皮初代培養系を用いました。すると、組織において上皮細胞シートに見られる大きな特徴の一つとしてあげられるPCPが、他の通常の培養細胞系では再現されないものの、気管上皮初代培養系では再現されました。PCPに従って、4つの配列状態の変化を経て、BBの規則的配列が成立する様子を世界に先駆けて明らかにすることができました(図2)。
多繊毛上皮の規則的繊毛配列のメカニズムについては、数理生物学的には、流体力学モデルで説明できることを示すことができました。すなわち、繊毛根元の「TJーアピカル複合体」のアピカル微小管の数や配列の変化から多繊毛上皮細胞アピカル表層のソフトマターの動態が変化することに起因して、BB配列が遷移することを数理的モデル化で説明することができました(図2)。

 gmail.com
gmail.com