大阪大学大学院 生命機能研究科

| 図1.核膜の構造 核膜は、核膜外膜・内膜、核膜内膜特異的タンパク質、核ラミナ、核膜孔複合体(NPC)など多くの機能部位からなる複雑な構造体です。核膜内膜タンパクのSUNタンパク質を介して、核ラミナと細胞骨格が繋がっています(詳細は「Chromatin Dynamics」 を参照)。 |
我々の研究室では、核膜の構造と機能の連関を明らかにするため、
1.核ー細胞質間の物質輸送の担い手:核膜孔複合体
2.核膜崩壊・形成過程
3.核膜による染色体機能制御機構
の3つに着目して研究を進めています。
1.核膜孔複合体
1-1.核膜孔複合体の構造
細胞核と細胞質は核膜によって隔離されています。核膜には核膜孔複合体(nuclear pore complex: NPC)と呼ばれる8回対称の筒状の構造体が、細胞質から核内まで貫通して埋まっています。NPCが形成する孔を通してRNAや蛋白質が核と細胞質の間で輸送されます。一方、最近ではNPCは核-細胞質間輸送を司る因子というだけでなく、エピジェネティクス・細胞分化・老化などにも関係する重要な因子としても注目されています。しかしNPCを構成する約30種類の蛋白質(ヌクレオポリン)がどのように機能をあるいは分担しあるいは連携して最終的な役割を果たすのかはわかっていません。私たちは、NPCを多様な細胞核機能に必須の機能構造体と捉え、特に以下のテーマについて研究を進めています。
①分化細胞の細胞核における核膜孔複合体の構造と機能
②分裂酵母のNPCの構造解明
③ヌクレオポリンの分子機能と細胞核機能の関係
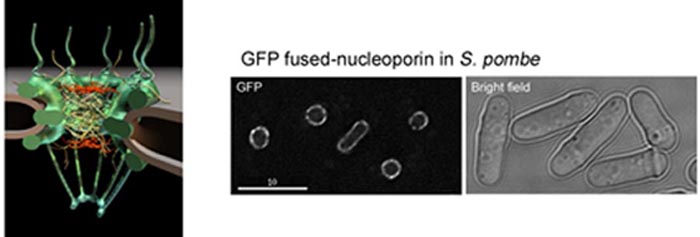
図2.核膜孔複合体(NPC)の構造と観察例
(左)核膜孔複合体の模式図 (右)分裂酵母におけるヌクレオポリン(GFP融合)の観察像
1-2.核膜孔複合体と細胞老化やガン化との関わり
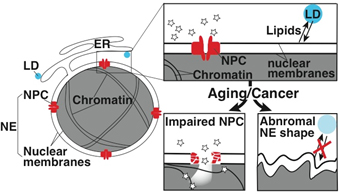 |
| 図3.Functions of the NPC and LD are crucial for maintenance of the NE integrity and homeostasis., respectively. (Left panels) Impaired NPC or disrupted lipid homeostasis causes loss of the NE integrity and abnormal NE shapes in the aging or cancer cells. |
2.核膜崩壊・形成過程(Virtual nuclear envelope breakdown)
高等真核生物の細胞では、分裂期に核膜が消失し(図3A:open mitosis)、核と細胞質が均一化します。一方、酵母に代表される菌類の多くは、核分裂過程で核膜は保持され(図3B:closed mitosis)、核蛋白質は全生活環を通して核 内に保たれます。分裂酵母も体細胞分裂期では核膜が消失しないことが知られていましたが、減数分裂の第二分裂では、核膜や核膜孔の構造を維持しながらも、機能的な「核膜崩壊」が起こり、核蛋白質が核の外へ流出することがわかりました(図3C:virtual nuclear envelope breakdown)。興味深いことに、このときにRanGAP1という核と細胞質の定義づけに重要な役割を果たす蛋白質の局在変化が起こります。核膜や核膜孔の構造を維持しながらも機能的には核膜消失と同じ効果がもたらされるというこの新しい分裂様式が、どのようなメカニズムで起きるのか、またそれがなぜ減数第二分裂でだけ起きるのかを追究していくことで、open mitosisとclosed mitosisの根源的な理解へとつながると考えられます。またそれによって核(膜)を持つ細胞の進化過程を知ることができるかもしれません。

図3.Open mitosis, Closed mitosisとVirtual nuclear envelope breakdown
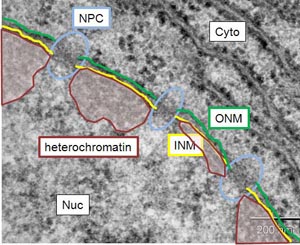 |
| 図4.核膜直下にはヘテロクロマチンと呼ばれる電子密度の濃い領域が存在する。 ONM:核膜外膜、INM:核膜内膜、NPC:核膜孔複合体、Nuc:核、Cyto:細胞質 |
我々は、核膜内膜特異的なタンパク質の1つであるラミンB受容体(Lamin B Receptor)がhistone H4のジメチル化された20番目のリジン残基を特異的に認識して染色体に結合し、転写抑制複合体を形成することを見出しました(Hirano Y. et al., J. Biol. Chem, (2012))。ラミンB受容体の変異体は、魚鱗症(皮膚が魚の鱗のようになる病気)やPelger-Huet anomaly(好中球の核分葉異常)の原因となること、これらの病気の細胞では染色体配置に異常が見られることが報告されており、我々の結果はLBRが染色体配置や転写状態を時空間的に制御することで、細胞分化の正常な進行に寄与していることを示唆しています。
さらに我々は、ヒトから酵母まで広く保存されている核膜内膜タンパク質の一つであるLem2(図5)が、栄養状態依存的にセントロメア領域に結合し、セントロメア周辺領域のヘテロクロマチン形成を促進することを見出しました(Tange Y. et al., Genes to Cells, gtc.12385 (2016))。
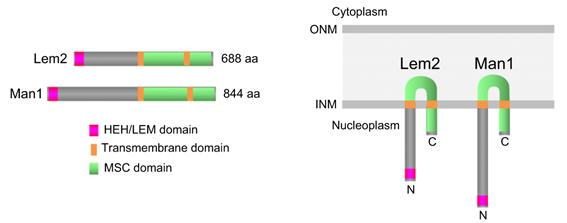
Lem2はLEMドメインタンパク質と呼ばれる核膜内膜タンパク質群に属し、N末端とC末端を核質側に突出した2回膜貫通型タンパク質。
また、Lem2の欠損では栄養状態依存的に染色体が不安定化することが分かっており、染色体構造の維持にも寄与していることが考えられます。これらより、Lem2は栄養状態の変化を検知して染色体構造や転写状態の制御を行い、環境変化に対応する役割があると考えられます(図6)。
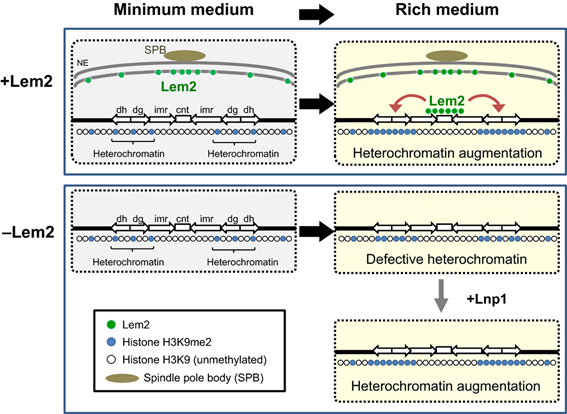
図6.我々のグループが明らかにしたLem2によるヘテロクロマチン形成機構。