細胞性粘菌Dictyostelium discoideumについて
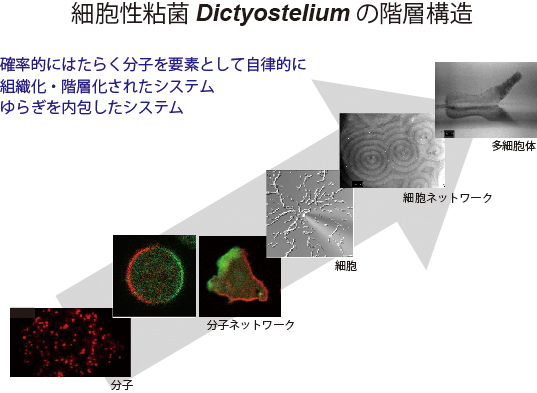
“分子機械の理解から分子機械システムの理解へ,さらには細胞からなるシステムとしての多細胞体への理解へと,生物の階層構造を飛躍することなく,また断絶を生むことなく理解できるのは,細胞性粘菌をおいて他にはないのではなかろうか.”(『モデル生物:細胞性粘菌』 p. 108)
細胞性粘菌Dictyostelium discoideum(和名キイロタマホコリカビ)の生活環の特徴
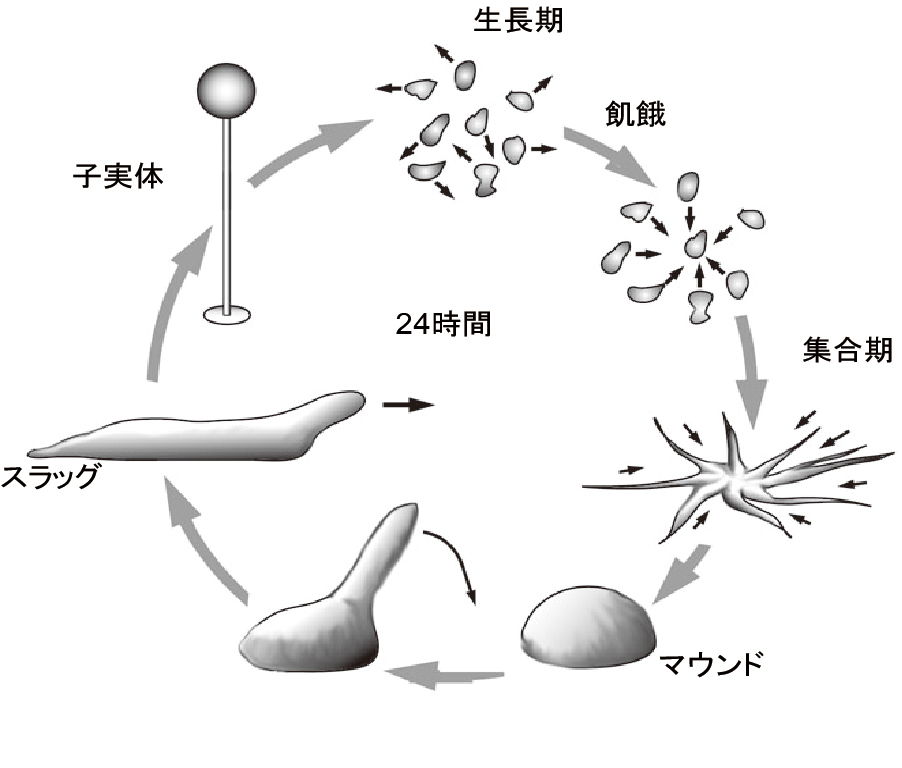 粘菌は土壌表層に広く分布しており,非常にしたたかな生活様式をもつユニークな生き物である.最終的な分化形態としては子実体と呼ばれる植物的な構造を形成する.子実体は胞子群とそれを支える細胞性の柄(死細胞)からなる.胞子は適当な条件下で発芽してアメーバ状の細胞となり,周囲の餌(大腸菌などのバクテリア)あるいは栄養豊富な培養液を取り込みながら増殖する.周囲の栄養源が枯渇して飢餓状態になると,細胞はやがて集合して多細胞体制を構築し,分化・パターン形成の方向に移行する.Dictyostelium discoideumでは,約10万個の細胞からなる半球状の集合塊(マウンド)の先頭部に乳頭突起(tip)が形成され,やがてナメクジ状の移動体となる.移動体には,運動方向に関して厳密な前後軸極性が存在するとともに,既に明瞭な分化パターンが認められる.すなわち,移動体の前部約1/4を占める細胞(予定柄細胞)は,子実体形成に際して,柄細胞に分化して死に,一方,後部約3/4の細胞(予定胞子細胞)は物理・化学的ストレスに対して抵抗性をもつ胞子に分化して生き延びる.これらのプロセスは無性的に進行する.(『モデル生物:細胞性粘菌』(前田靖男氏編)から引用させていただきました.pp. i-ii)
粘菌は土壌表層に広く分布しており,非常にしたたかな生活様式をもつユニークな生き物である.最終的な分化形態としては子実体と呼ばれる植物的な構造を形成する.子実体は胞子群とそれを支える細胞性の柄(死細胞)からなる.胞子は適当な条件下で発芽してアメーバ状の細胞となり,周囲の餌(大腸菌などのバクテリア)あるいは栄養豊富な培養液を取り込みながら増殖する.周囲の栄養源が枯渇して飢餓状態になると,細胞はやがて集合して多細胞体制を構築し,分化・パターン形成の方向に移行する.Dictyostelium discoideumでは,約10万個の細胞からなる半球状の集合塊(マウンド)の先頭部に乳頭突起(tip)が形成され,やがてナメクジ状の移動体となる.移動体には,運動方向に関して厳密な前後軸極性が存在するとともに,既に明瞭な分化パターンが認められる.すなわち,移動体の前部約1/4を占める細胞(予定柄細胞)は,子実体形成に際して,柄細胞に分化して死に,一方,後部約3/4の細胞(予定胞子細胞)は物理・化学的ストレスに対して抵抗性をもつ胞子に分化して生き延びる.これらのプロセスは無性的に進行する.(『モデル生物:細胞性粘菌』(前田靖男氏編)から引用させていただきました.pp. i-ii)
細胞性粘菌Dictyostelium discoideumにみられる走化性・走電性・走光性・走熱性などの走性行動
細胞走性とは,細胞が環境にある化学物質や光,温度など空間勾配を認識して,方向性のある移動運動を行う性質である.化学物質に対する応答は走化性応答 — 他, 走光性,走熱性,走電性など — と呼ばれる.バクテリアを捕食して増殖する生長期には,バクテリアが分泌する葉酸に対して走化性応答を示す.バクテリアを食べ尽くして飢餓状態になると,細胞自ら産生するサイクリックAMP(cAMP)に対して走化性を示すようになり,細胞が集合して多細胞体のスラッグをつくる.ナメクジのような形をしたスラッグは,光に向った走光性や増殖期の温度に向った走熱性を示す.また,生理的な意義は未確認であるが,生活環の各ステージで電場に対する走電性を示すことができる.
{kind=link}
cAMPに対する走化性応答と細胞内1分子イメージング法の開発に取り組んだきっかけ
真核生物の走化性の分子メカニズムについては,細胞性粘菌でもっとも理解が進んでおり, 走化性研究のモデル生物となっている.特に,集合期におけるcAMPに対する走化性の分子論的理解が進んでいる. 粘菌細胞は僅かなcAMPの濃度差(平均濃度1nMで濃度勾配2〜5%程度)に対して走化性を示すことができる. このとき細胞に結合する誘引物質の分子数を計算してみると,計算上は,細胞全体で約300分子の誘引物質が受容体に結合し, 細胞の前半分と後半分で4〜10分子程度の差があると,細胞は濃度勾配の方向を認識できることになる.誘引物質と受容体の 結合・解離反応は確率的なので,活性化した受容体の数は時間的に一定せず,平均値の周りにゆらぐはずである.このとき の平均値を誘引物質濃度に対応したシグナルと考えると,そのゆらぎは濃度情報の読み取り誤差(ノイズ)と言える.この ノイズの量は数十分子程度となり,細胞に濃度勾配の情報を与える誘引物質結合数の前後差よりも大きいぐらいである. 『ノイズに満ちたシグナルから微弱な濃度勾配の情報を取り出し,安定した走化性応答を実現するために,細胞内では どのようなタイプの反応が用いられ,それらを組み合わせた反応経路はどのようにデザインされているのか?』. 細胞性粘菌の走化性シグナル伝達系の1分子計測法を開発しようと考えたのは,この問いを解きたいと思ったからである. こうした研究を始めた頃(1998年),佐甲靖志氏(現在理化学研究所)によって細胞内で1分子を見る技術がまさに誕生 しつつあった.細胞内1分子計測法を適用して,細胞内のシグナルを分子の数として数えることができれば, 実際にシグナルがどれほどノイジーなのかが分かるだろうと考え、この研究を始めた.