【回答】
★グループ1 **************************************************************************************
現存するモータータンパクの作動原理を調べるのではなく、“改良型モータータンパクを実際に創り出し、天然タンパクと比 較することで、その作動原理に迫る”という構成的アプローチをとる。
具体的には、アクチン-ミオシンの系で高速走行型ミオシンを進化工学的手法で作り出 す。一般的変異型解析と違うのは、既知構造情報にとらわれないランダム変異ライブラリーを扱うことで、変異体の探索ができるところである。
<方法 : 進化工学的手法>
ランダム変異ライブラリーからリボソームディスプレイ法によ りgenotype- phenotype linkage(遺伝子型と表現型を対応付けた複合体)を実現する。リボ ソームディスプレイ法とは、mRNAの終止コドンを取り除くことでリボソームの遊離を抑え、遺伝 子型としてのmRNAと表現型としてのタンパク質とリボソームの複合体を形成させ る方法である。
mRNA リボソーム ミオシン

このようにして作製した無数の変異型ミオシンgenotype-phenotype linkageを アクチンレール上で競争させることで、高速走行型ミオシンを選択することができる。
こうして取得した高速走行型ミオシンに対して、変位解析、ATPase活性測定、構造解析、シミュレーションなどの解析を行う。
特に、これらの変異体のうち、ATPaseあたりの動きも変化しているものの構造を解析・比較すれば、 力学的性質のシミュレーションも可能である。
★グループ2 **************************************************************************************
① ア クチン-ミオシンの系を1分子レベルで解析することは、アクチンのステップ的な動きな ど様々な情報を得ることに成功しているが、1分子観察ではシグナルが弱すぎてどうしても観察できない部分 も多い。そこでここでは、多分子のアクチン-ミオシン系で、ATP加水分解にともなうミオシンの挙動を詳細に観察する方法を考 える。そこで重要になるのが観察対象になるミオシン全体の動きを同期させなければならないということである。そのための方法として、各ミオシンの頭部を化 学的に架橋して、ミオシンを縦横に整列させ、さらにそのミオシンの隊列をアクチンのシートで挟むという下図のような構造を考える。
ミオシンを30人31脚&ムカデ競走状態にす
る アクチンシート
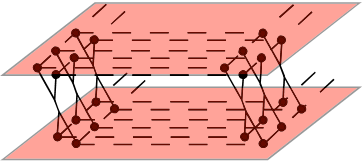 このよ
うにしてミオシンの動きを同期させる。しかし実際に人間が30人31脚またはムカデ競走をする場合を考えれば予想できるように、
すぐに動きがずれてつまずいてしまうだろう。おそらく、きちんと同期するのは最初の2・3歩くらいであり、その時が解析すべき対象となる。したがっ
て、上の図のようなシート状の構造をあらかじめ作った上でATPを加え、その直後を解析しなければならない。
このよ
うにしてミオシンの動きを同期させる。しかし実際に人間が30人31脚またはムカデ競走をする場合を考えれば予想できるように、
すぐに動きがずれてつまずいてしまうだろう。おそらく、きちんと同期するのは最初の2・3歩くらいであり、その時が解析すべき対象となる。したがっ
て、上の図のようなシート状の構造をあらかじめ作った上でATPを加え、その直後を解析しなければならない。
解析の手法はX線回折とFRET法を用い、1分子では捕えられなかった構造変化を見る。動きが速くて検出 できないときは、温度を下げたり、シートを引っ張ったり、変異を入れたミオシンを用いるなどする。
② F1 ATPaseを 人為的に回転させてATPを合成させたように、ミオシンにADPとPiを加え、ミオシンの頭部構造を引っ張ったりゆがめたり、無理 やり何段階かの変化を加え、ATPが合成される経路を模索する。ATPが合成される構造変化が見つかったら、その逆の反応経路が、ATPが加水分解されてADPとPiができる過程であり、すなわちATPのエネルギーがミオシンの構造変化に変換される過程である。
③ ミ オシンにはATP加水分解過程において複数の中間体が存在し、それらのいくつ かについてはX線結晶構造がわかっている。そこで、わかっている中間体の結 晶構造情報から、それらのエネルギー状態を予測し、さらにミオシンのモーター部位などでの構造変化にかかるエネルギーを推定する。こうして各中間体のとり うる構造変化とそれに必要なエネルギーを算出して、1分子のATPが加水分解されて生じるエネルギーがミオシンの構造変化にど のように分配されるのかを考える。
★グループ3 **************************************************************************************
アクチン-ミオシンの系で
そもそもミオシンのステップ的動きにエネルギー変換メカニズ ムは必要なのだろうか。ミオシンのステップ的動きをよく観察してみると、必ずしも常に一定方向に動いているわけではないことが分かる。このことは、個々の ステップはブラウン運動によるものでエネルギーを必要としないという印象を与える。そこで、運動そのものにはエネルギーは必要ではなく、ミオシンがブラウ ン運動でゴール位置に到達したときにセンサーが働いてミオシンがキャッチされると考えることはできないだろうか。つまりエネルギー変換の過程は、ミオシン がある方向に数ステップ進んだことを検知するセンサーのオン・オフにあると考えるわけである。
それでは、上記の可能性と従来の考え、すなわちATP加水分解エネルギーが生体分子の構造変化を引き起こし、それ がステップ的運動につながるとする考えを区別しなければならないが、そのためには、ミオシンのアームの部分の固さを変異の導入などにより変えてやるとよい のではないだろうか。
鞭毛モーターの系で
鞭毛モーターでは、細胞内外のプロトン濃度勾配に従って流入 するプロトンが、その流入過程で、あるタンパク質との結合・解離反応を起こし、そのタンパク質の構造変化を引き起こすことでステップ状の回転運動を生み出 しているとするモデルがある。したがって、鞭毛のエネルギー変換メカニズムを解明するためには、プロトンとの結合・解離によって変化するタンパク質の構造 解析が重要になる。
そこでX線結晶解析を行うが、結晶構造はあくまでもタンパク質の静止 した情報しかもたらさないので、構造変化の前と後、また中間状態の構造も解析しなくてはならない。問題のタンパク質はプロトンの結合・解離によって構造を 変化させるので、様々なpHにおいて結晶を作成し、それぞれの解析情報を統合すれば、構 造変化の一連の流れを予測することができるだろう。
★グループ4 **************************************************************************************
分子モーターでのエネルギー変換メカニズムとは、化学エネル ギーが構造の変化という力学エネルギーに変換されるメカニズムである。より詳細には、たとえばミオシンの場合を考えると、ATPの高エネルギー結合の開裂がATPase中のヌクレオチド結合部位のオングストロームレベルの構造変 化を引き起こし、それが計測可能なミオシンの大きな構造変化やステップ的運動につながるという一連の過程である。この流れを順を追って眺めることができれ ば、エネルギーが変換される瞬間をとらえることができるだろう。
ところが、問題を複雑にしているのは、1)Piの放出の速さ(~ms)と、2)それを検出する手法がないこと、また3)Pi放出により引き起こされたオングストロームレベルの構造変化 をリアルタイムでとらえる方法がないことである。各問題点は以下の通り。
1) Pi結合タンパク質を使用したストップドフロー計測は~1.5msのデッドタイム。
2) そ のため、それ以上高速の現象をとらえることができない。
3) Pi放出前後のオングストロームレベルの残基の構造変化は、ヌク レオチドアナログを用いたX線結晶構造解析により、それなりの予想がされているが、時間 を追ってモニターする方法がない。
これらの問題を解決する方法として、Luca Turinの“分子振動共鳴仮説”や、少し脱線して車のロータリーエンジンとレシプロカルエン ジンのエネルギー効率の違いが生み出される理由、ATPの高エネルギー結合とは何か?など様々な意見交換が行われ た。
そのなかで、Piの放出が見えないのなら、こちらから積極的に別の方法で(エ ネルギーの入力は、化学エネルギーであろうと力学エネルギーであろうとかまわないはずなので)同じ現象を引き起こせばよいのではないかというアイデアにま とまった
具体的には、Pi放出で引き起こされると予想されるオングストロームレベルの 構造変化を、DNAのひもを取り付けた分子モーターを一分子操作によってマリオ ネットのように操ることで再現してやるというものである。名付けて、“分子モーター・マリオネット計画”である。
例)
ミオシン Pi 光ピンセット DNA 大きな構造変化 Åの構造変化![]()
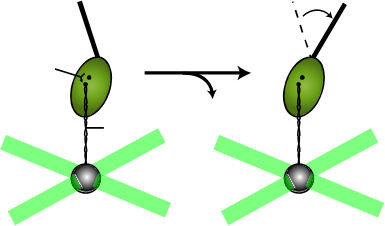
この実験で、化学エネルギーによりもたらされると考えられる 微妙な構造変化を複数試し、自然な力学的アウトプットを示すオングストローム構造変化を特定する。このようにして特定した小さな構造変化は、エネルギー変 換の核となるPi放出をミミックしたものであると考えられる。
F1-ATPaseの場合は順番に引っ張ることでATPなしで回転させることができるかもしれないし、ミオシンⅤの 場合には右足と左足を交互に引っ張ることでアクチンの上を歩行させることもできるかもしれない。