
-
565-0871
大阪府吹田市山田丘1-3
大阪大学
生命機能研究科
発生遺伝学グループ
06-6879-7994
体軸の始まり(前後軸形成からのアプローチ)
 ショウジョウバエを含む多くの生物は、すでに受精前の卵子の段階で前後方向が決定しています。対して、私たち人やマウスといった哺乳類は、受精卵を2つに分けてもそれぞれが正常な胎児へと発生するという高い適応能力を持っていることから、卵子や受精卵の時期においては非対称性を獲得しておらず、その後の発生過程で「非対称性の起源」というイベントを経て、非対称性を獲得すると考えられています。 私達は、体軸の中でも一番早期に決まる前後軸に着目し、初期メカニズムをさかのぼることで「非対称性の起源」に迫ろうというストラテジーのもと研究を行っています。
ショウジョウバエを含む多くの生物は、すでに受精前の卵子の段階で前後方向が決定しています。対して、私たち人やマウスといった哺乳類は、受精卵を2つに分けてもそれぞれが正常な胎児へと発生するという高い適応能力を持っていることから、卵子や受精卵の時期においては非対称性を獲得しておらず、その後の発生過程で「非対称性の起源」というイベントを経て、非対称性を獲得すると考えられています。 私達は、体軸の中でも一番早期に決まる前後軸に着目し、初期メカニズムをさかのぼることで「非対称性の起源」に迫ろうというストラテジーのもと研究を行っています。
 これまでに、前後軸ができなくなるノックアウトマウスの解析(Yamamoto et al., Genes&Development 2001)を皮切りに、前後軸方向は受精後5日で決まっていること (Yamamoto et al., Nature 2004)。さらにさかのぼって、受精後4日目で決まっていることを明らかにしました (Takaoka et al., Dev. Cell 2006) (Takaoka et al., Nature cell biology 2011)。現在、さらに早期にあるであろう「体軸の始まり」へ挑戦中です。
これまでに、前後軸ができなくなるノックアウトマウスの解析(Yamamoto et al., Genes&Development 2001)を皮切りに、前後軸方向は受精後5日で決まっていること (Yamamoto et al., Nature 2004)。さらにさかのぼって、受精後4日目で決まっていることを明らかにしました (Takaoka et al., Dev. Cell 2006) (Takaoka et al., Nature cell biology 2011)。現在、さらに早期にあるであろう「体軸の始まり」へ挑戦中です。

図1 従来の頭尾(前後)軸形成モデルと本研究から明らかになった新モデル
- 従来のモデル(上段):
- DVEは受精後5日で形成され(緑)、将来の頭側へ移動し、AVEと名前を変え、6日胚では頭を誘導します。
- 新モデル(下段):
- 頭側を誘導するDVEは少なくとも4日胚で形成される(緑)。DVE/AVE発現遺伝子(例:Lefty1)は、DVEが将来の頭側へ移動すると共に発現が消失し(緑白抜き)、遠位側では新たに発現を開始します(ピンク)。
~最近の知見より~
(Takaoka et al., Nature cell biology 2011)
マウス胚における前後軸は受精後5日目から6日目にかけて決まっていると考えられてきました(図)。具体的には、受精後5日胚で胎盤に対して遠位側にDVEと呼ばれる胚の頭側(前側)を決める特殊な細胞群が現れ、その後、DVE細胞が頭側へ移動し、AVEと名前を変えて頭部誘導シグナルを分泌することにより、AVEから近い位置にある胚体部分(将来体になる部分)は頭側に、遠い位置にある胚体部分は尾側になるとされてきました。
しかし私達は、遺伝学的な手法を用いて細胞の運命・由来を調べるとともに、蛍光タンパク質を用いてリアルタイムで細胞の挙動を観察することで、従来の知見を覆し、新たな前後軸形成モデルを提唱しました。まず、①DVEになるべき細胞は、遅くとも受精後4日目にすでに決定されていること、②同じ細胞と考えられてきたDVEとAVEは、実は異なる由来の細胞であること、③DVEの役割は、自らが頭側へ移動することにより、遅れて生じるAVEを頭側へガイドすることなどが明らかになりました(図1)。このように、常識化されていた考えにとらわれずに、細胞の由来・挙動という新たな観点から観察することで、新たな発見につながりました。 今後は、さらに「体の非対称性の起源」へ迫るために、受精後4日目よりも遡って研究を行います。特に、DVEという前後方向を決める特別な細胞がどのようにして、均一な極性のない細胞から選択されるのかという疑問を明らかにしたいと考えています。
ES細胞やiPS細胞から複雑な構造を持つ臓器を作るためには、目的の臓器の細胞を誘導するだけではなく、細胞集団へ正確な位置情報を与え、高度な構造を持たせることも重要です。本研究から得られた知見は、体の非対称性の起源という発生生物学の命題に迫るとともに、将来の再生医療の基盤になるものと期待されます。
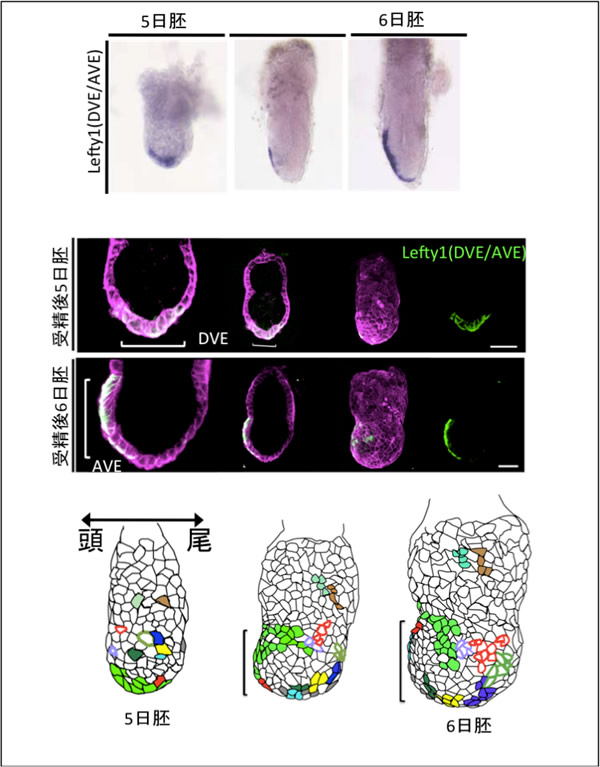
図2 蛍光たんぱく質を用いて明らかになった細胞分布図
- 上段: 5日胚ではDVEで、6日胚ではAVEに発現している。
- 中段: VEの細胞膜を赤色蛍光たんぱく質、DVE/AVEの細胞膜を緑色蛍光たんぱく質で標識しました。経時的に観察することで、1細胞レベルの挙動を知ることができます。
- 下段: 5日から6日胚の細胞分布図。DVE(緑)の細胞とAVE(赤、水、灰色)の細胞は異なる細胞でした。
